El tejido nervioso se desarrolla al comienzo
de la tercera semana de vida intrauterina. Se origina de la capa germinativa
denominada ectodermo específicamente de su parte dorsal.
Primero se forma un engrosamiento que se
denomina placa neural y posteriormente se invagina para dar origen al canal
neural. Este canal se encuentra limitado por dos crestas laterales que corren
del extremo oral al extremo anal del embrión. La transformación del canal en
tubo inicia en la parte media del embrión hacia sus extremos. El tubo neural no
alcanza a cerrarse del todo, dejando en sus extremos dos poros denominados el
neuroporo anterior y el neuroporo posterior. El neuroporo anterior o cefálico,
mediante el desarrollo del tejido nervioso se oblitera por la lámina terminalis
que en el adulto toma el nombre de lámina supra óptica, por quedar situada
inmediatamente por encima del quiasma óptico. El neuroporo posterior se
oblitera por tejido nervioso, no teniendo interés alguno en el adulto.
La mayor parte del sistema nervioso deriva del
tubo neural y de las crestas neurales llamadas también crestas ganglionares,
otras porciones derivan de las placodas ectodérmicas.


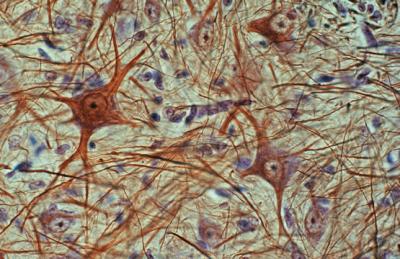
Histología:
Es un tejido formado principalmente por dos
tipos celulares: neuronas y glía, y cuya misión es
recibir información del medio externo e interno, procesarla y
desencadenar una respuesta. Es también el responsable de controlar numerosas
funciones vitales como la respiración, digestión, bombeo sanguíneo del corazón,
regular el flujo sanguíneo, control del sistema endocrino, etcétera.
Las células del
sistema nervioso se agrupan para formar dos estructuras: el sistema
nervioso central que incluye el encéfalo y la médula espinal, y
el sistema nervioso periférico, formado por ganglios, nervios y neuronas
diseminados por el organismo.
El tejido nervioso dispone tan sólo de dos
tipos de células específicas: las células
nerviosas o neuronas y
las células gliales o neuróglicas. Sólo las neuronas
intervienen de forma directa en los procesos informativos. Las células gliales
se vinculan con los vasos sanguíneos y el tejido conectivo, los cuales
pertenecen a la parte inespecífica del sistema nervioso y sus funciones son
exclusivamente metabólicas o mecánicas.
En el sistema nervioso
central hay zonas ricas en cuerpos celulares de neuronas y glía que se denomina
generalmente sustancia gris, porque tienen un color gris en el tejido
fresco, mientras que las zonas ricas en axones mielínicos pero con pocos
cuerpos celulares se denominan sustancia blanca. La sustancia blanca es
una zona de tractos de fibras. En el encéfalo, la sustancia gris es normalmente
superficial, mientras que en la médula espinal es al contrario.
La función esencial del tejido nervioso es la
comunicación, que depende de la capacidad de las células nerviosas para recibir
estímulos (excitabilidad) y la propiedad de transmitir la excitación resultante
desde un punto hasta otro (conductividad). En consecuencia, relaciona los
ambientes interno y externo, que se analizan e integran para producir
respuestas coordinadas. El tejido nervioso posee una organización estructural y
complejidad funcional tan específicas, que se lo conoce habitualmente
como sistema nervioso